The Plesiosaur Mystery Bone
or, Why Do Marine Reptiles Keep Evolving Extra Bones?
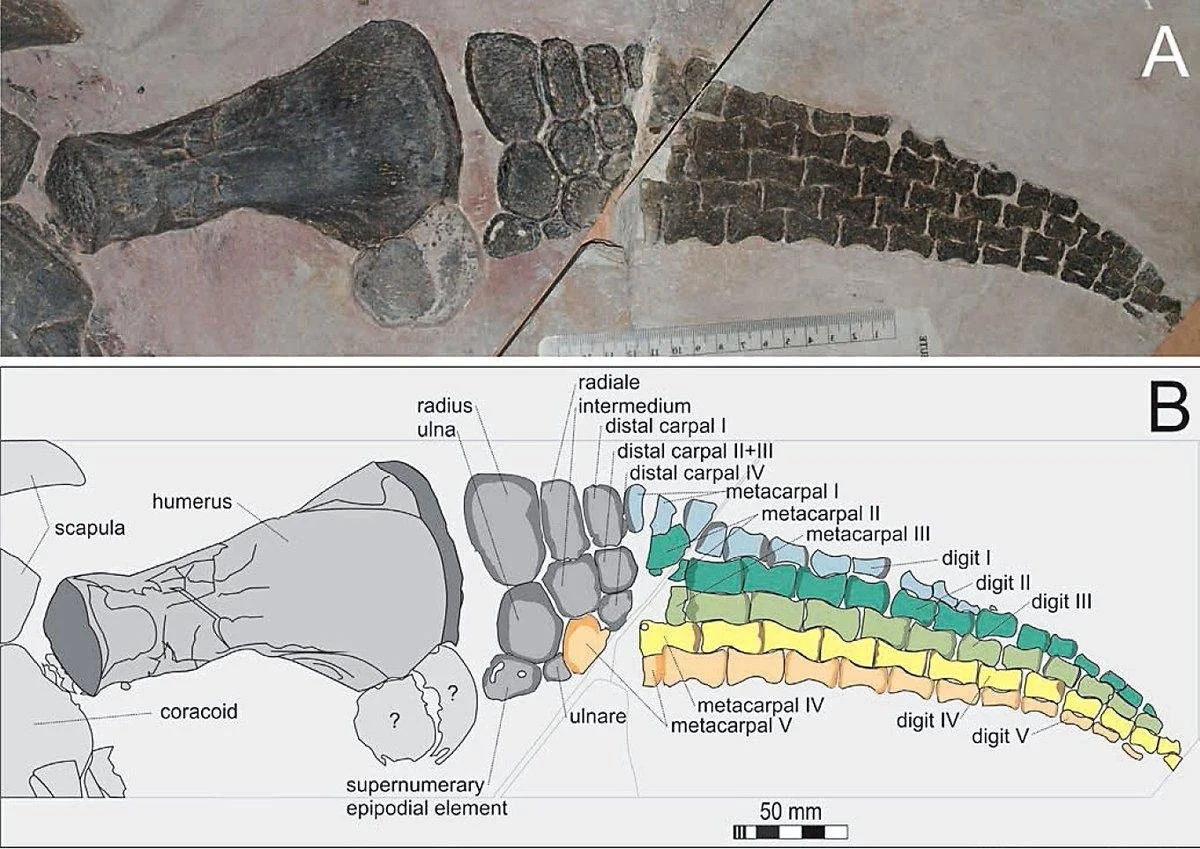
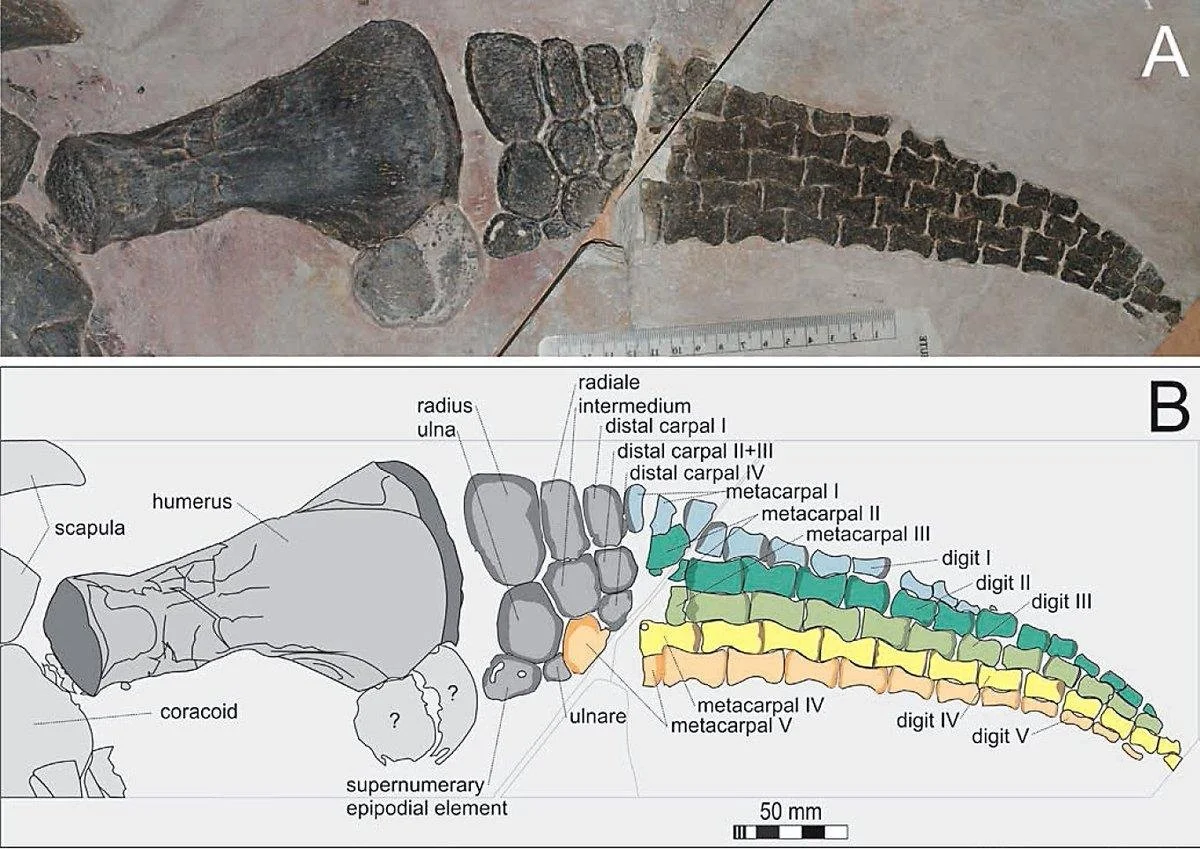
Flipper of the polycotylid plesiosaur Mauriciosaurus.
One of the craziest facts I learned studying marine reptiles is that certain plesiosaurs- polycotylids and leptocleidids- have an extra bone in the forearm and lower limb, which as far as I know isn't seen in any other animals. Most tetrapods have the standard "one-two-many" structure of the limbs: one bone in the proximal limb (humerus or femur) then two (radius+ulna or tibia+fibula) then lots of little bones (carpals/tarsals, metapodials, phalanges). Leptocleidids and polycotylids have a "one-three-many" structure: three bones contact the end of the humerus and femur.
So we have this strange bone lying between the humerus and the wrist, (and another between the femur and the ankle), and we don’t actually know what these bones are. Most bones have an anatomical term (humerus, radius, ulna, metacarpals, etc.) and this one doesn’t— it doesn’t even have a name. But presumably given its position in the forelimb/hindlimb it’s a serial homologue of the other two- basically, a duplicate ulna / fibula.

The Kem Kem leptocleidid was a small plesiosaur that lived in freshwater along with the giant Spinosaurus
I encountered this mystery bone working on plesiosaurs from the Kem Kem beds of Morocco. It turns out plesiosaurs are fairly common in the Kem Kem, even though it’s a freshwater ecosystem, but nobody noticed because nobody was looking for them.
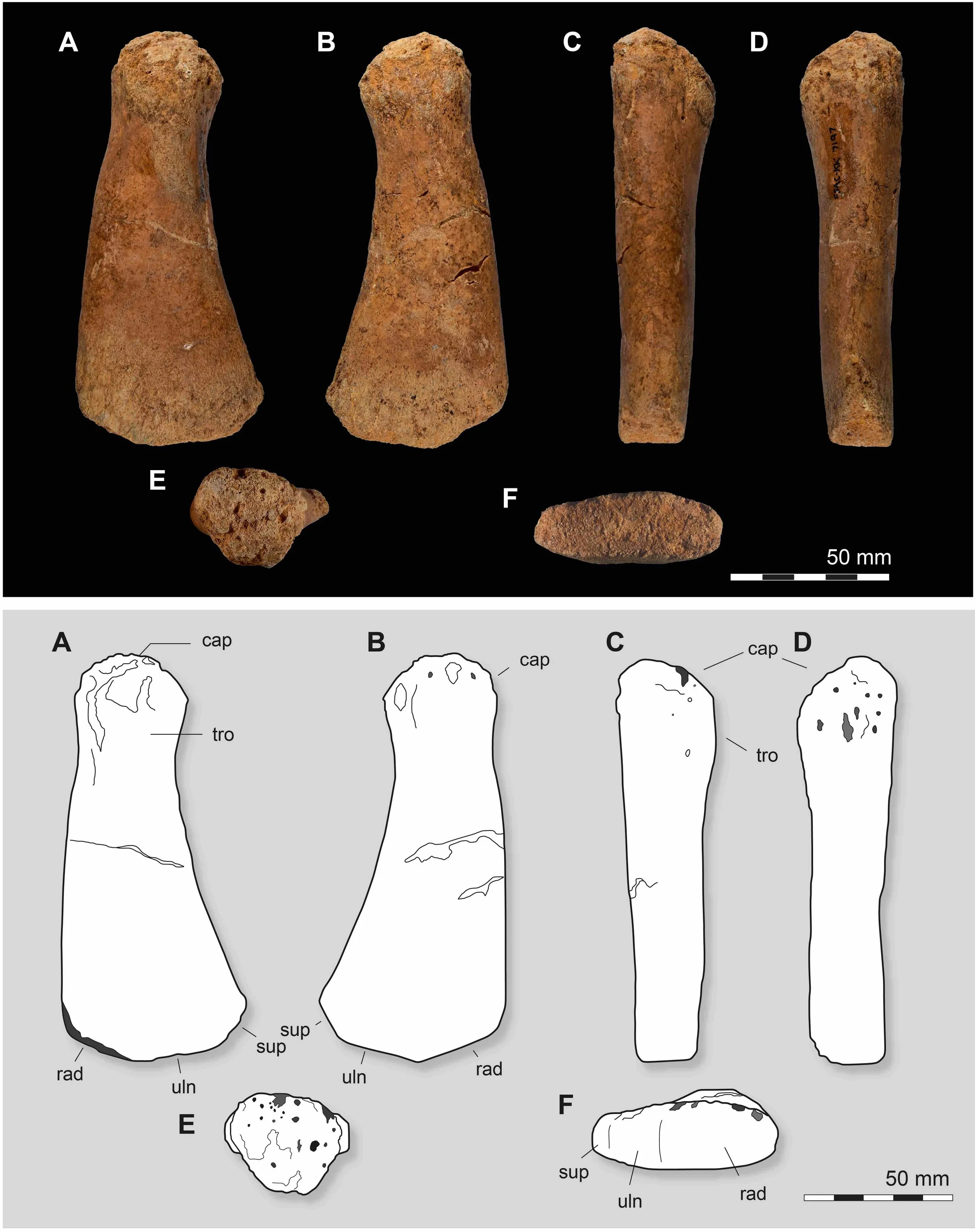
Kem Kem leptocleidid humerus. Rad = radius, uln = ulna; the contact for the Mystery Bone is ‘sup’ or “supernumerary epipodial element” which is a fancy way of saying “extra bone”.
I stumbled on this problem when I found a small plesiosaur humerus in a rock shop in Morocco. In the process we spent a lot of time comparing this little arm bone to lots of other plesiosaur arm bones, and it seemed we most likely had a leptocleidid.
One unusual feature that suggested a leptocleidid was this extra articulation at the end of the humerus. This isn’t a general plesiosaur thing, but specific to the family Leptocleididae and the related Polycotylidae. Animals like Plesiosaurus, Pliosaurus and Elasmosaurus lack it. Even among plesiosaurs, which are pretty weird, this anatomy is strange.
My best guess is that the developmental pattern that creates the ulna in the forelimb and the fibula in the hindlimb has somehow duplicated these elements- it’s what you call a serial homologue of the other forearm bones, the same bone repeated in a different place. This hypothesis might seem unlikely because nothing else has ever done this, but I don’t have another explanation. None of the wrist or ankle bones seem to be missing, so it can’t be a bone from the wrist /ankle that’s been shifted proximally. It’s not impossible it’s an extra carpal/tarsal but this would also be highly unusual.
In short: it’s a “supernumerary epipodial element” which is a fancy way of saying it’s an extra leg bone we don’t even have a name for…
A Mystery Bone.
Why does this thing exist? Well, here’s where things get really interesting, and even murkier.
Although these extra arm/leg bones are unique, they’re part of a broader pattern of extra bones evolving in the limbs of marine tetrapods like plesiosaurs, ichthyosaurs, mosasaurs, and whales.
Take another look at the polycotylid flipper. The most obvious feature isn’t that one little extra bone. It’s the insane number of phalanges- finger bones- and knuckles between them. We have two bones in our thumb, and three in the index, middle, ring and pinky fingers- a 2-3-3-3-3 arrangement.
The polycotylid Mauriciosaurus has at least 8 bones in the thumb, 13 in the index finger, and at least 11 or 12 in the remaining digits. Between the two hands, that’s over 100 knuckles. The precise number varies from species to species but all plesiosaurs show this— a radical departure from other reptiles like lizards and dinosaurs. The technical term is “hyperphalangy”, a fancy way of saying, “extra finger bones”.
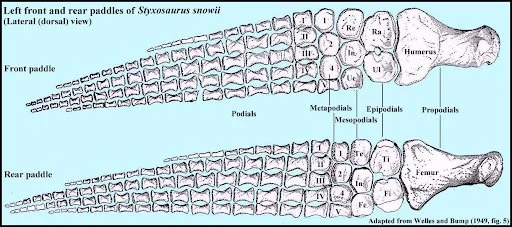
Forelimb (top) and hindlimb (below) of the elasmosaur Styxosaurus

The Jurassic Plesiosaurus, showing lots of finger and toe bones.
Hyperphalangy isn’t unique to plesiosaurs. Mosasaurs also add finger and toe bones, especially in later, more advanced mosasaurs like Mosasaurus and Plotosaurus.

Mosasaur flipper with extra finger bones.

Mosasaurus, with even more bones in the hand
We also see an increase in the number of finger bones in ichthyosaurs- some of the fingers of Ichthyosaurus have more than 20 phalanges.

Ichthyosaurus forelimb, showing extra finger bones.
And we also see extra finger bones in whales. In a dolphin, the index and middle fingers have more phalanges, the thumb is unchanged, the ring and pinky actually have fewer bones- but overall there’s more bones.

Dolphin flippers, showing extra bones in the index and middle finger
Ichthyosaurs show another bizarre pattern. Some increase the number of fingers- a mutation known as polydactyly.
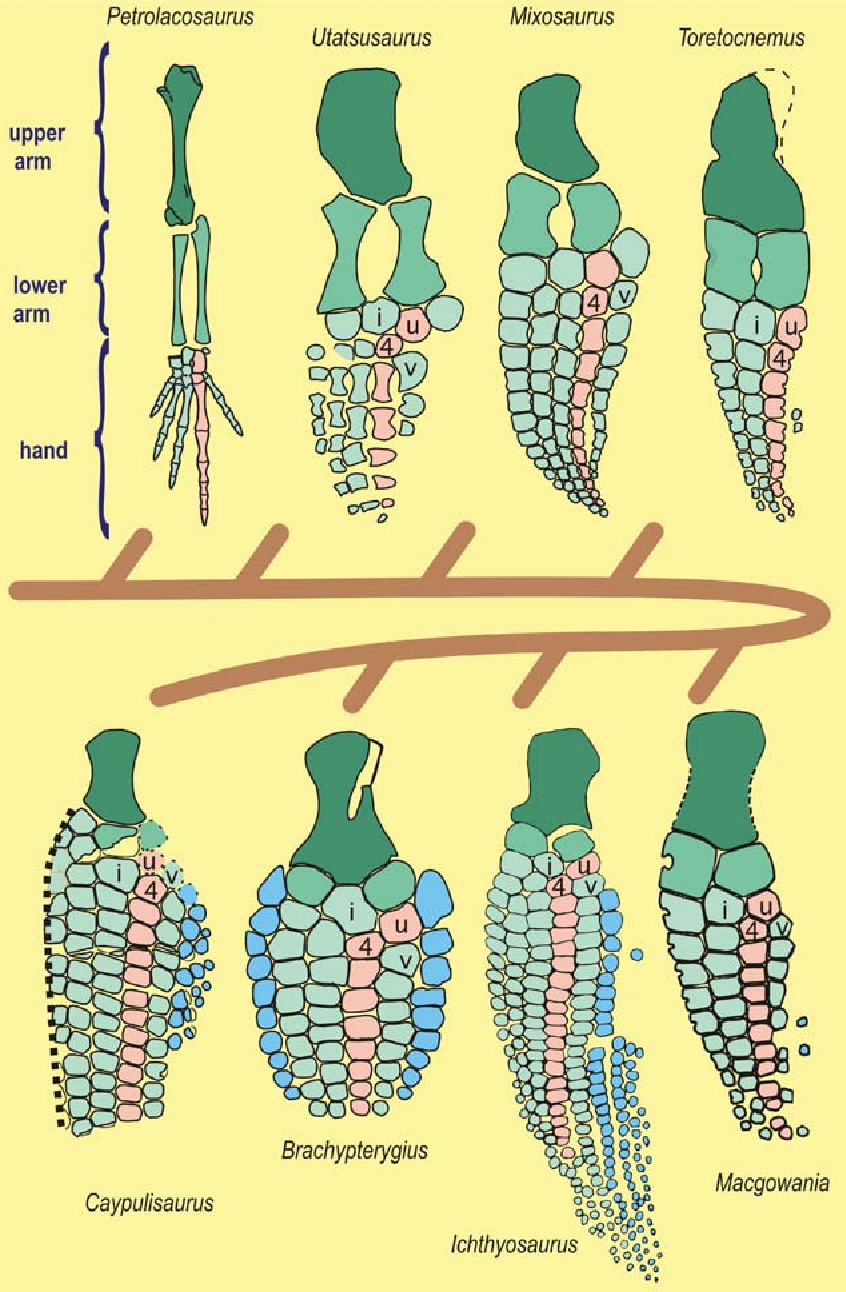
Ichthyosaur flipper evolution
A number of species like Ichthyosaurus and Brachypterygius evolve a sixth finger, a pinky to the pinky. Then something truly bizarre happens. This extra pinky forks, and forks, and one of the forked digits forks…
In certain animals this kind of thing is fairly common as a developmental anomaly. Cats often have extra digits, and sometimes these form by forking. But in plesiosaurs it’s not a developmental defect, it’s a feature, preserved by natural selection. This pattern isn’t universal. Some plesiosaurs actually reduce the number of fingers, but overall they gain bones rather than lose them.
Which raises a question: why?
The common theme here is a shift from the use of the limbs as weight-bearing structures on land to fins in water. Something about that shift- from foot to fin, or from land to the weightlessness of water, or both- may release a structural constraint on skeleton evolution, or favor a different design.
A couple observations fit this hypothesis.
First, if we look at animals that make a partial transition to the sea, they don’t show extra bones. Sea turtles, seals, sea lions, and penguins (which come back to land to breed) retain a fairly typical forelimb and hindlimb skeleton. They use their limbs as flippers and fins, but also to walk, and so retain a more typical tetrapod limb skeleton. The constraints of weight-bearing on land seem to prevent the weirdness you evolve in plesiosaurs, mosasaurs, and ichthyosaurs.
Sea lion flipper
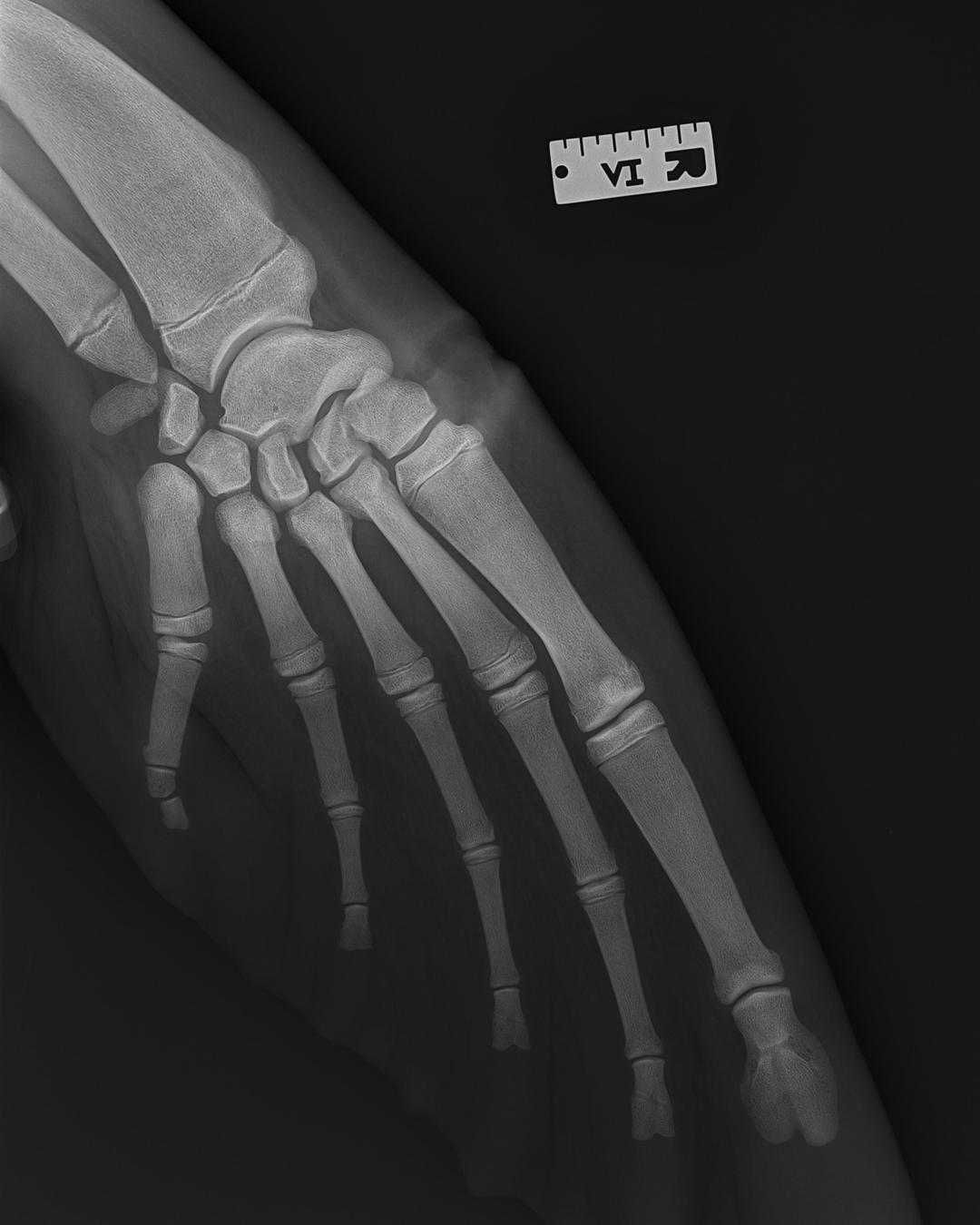
Another observation fitting this pattern is that early tetrapods- animals like Ichthyostega and Acanthostega, which used their limbs to swim and creep along riverbeds rather than to crawl on land- had a large number of fingers and finger bones- eight toes and multiple bones per toe.
After tetrapods make a complete transition to land, they decrease the number of fingers and finger bones… a trend that is reversed when tetrapods go back to the sea.
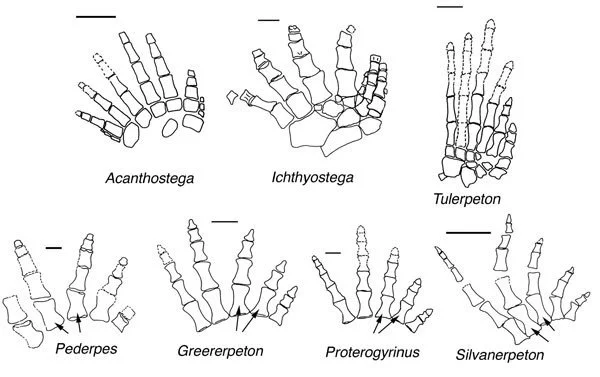
Early tetrapod foot skeleton
Exactly what drives this is unclear.
One hypothesis is that the reduction in number of bones is more structurally efficient— fewer bones are, for a given amount of material, stronger than many bones. Several observations fit this hypothesis.
Strikingly, many fast-moving animals have reduced the number of bones in the hands or feet. Theropod dinosaurs reduced their number of toes from five to four. Some fast runners like ornithomimids and avimimids went further and reduced their toes down to just three. Ostriches have only two toes.
Among mammals, horses have only one toe, the extinct litopterns of South America evolved something similar.
Since the foot is swung quickly during locomotion, reducing mass will (like a lightweight pair of running shoes) allow you to swing your foot faster and with less energy. Getting rid of toes and bones may make walking and running faster and more efficient.

Bird hand (here, a white ibis, Eudocimus) showing the reduced number of finger bones and the fusion of the hand and distal wrist bones.
The place where weight reduction will be most important, obviously, is flight. Here, every milligram of bone and tissue matters; since you need to generate more lift to carry that weight.
And we see something very curious: birds and pterosaurs dramatically reduce the number of bones in the limbs and skeleton by eliminating and fusing bones. Pterosaurs eliminate the pinky finger, and fuse their carpal bones so that they have just two elements in the wrist. Birds reduce the number of bones in the third finger, and fuse the hand and distal wrist bones.
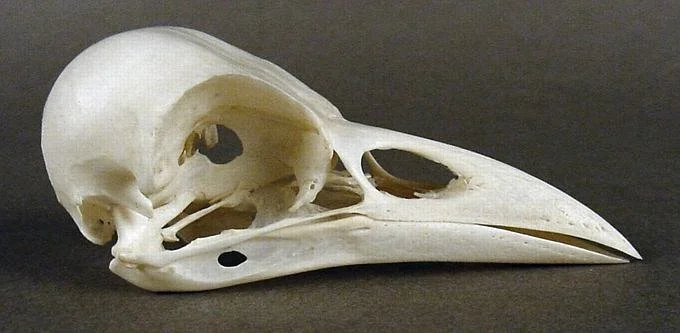
Almost all the bones of the bird skull and jaw are fused: functionally there is one big “Skull Bone”, a “Jaw Bone” and a few free elements like the quadrates and pterygoid.
Birds also fuse their skull bones together, eliminating the complex sutures and joints to create a single structure incorporating almost all the bones of the skull, and fusing the lower jaw bones together. So do pterosaurs.
Both pterosaurs and birds fuse the vertebrae of the back together into a structure called the notarium. And they fuse dorsal, sacral and caudal vertebrae and hip bones into a compound bone called the synsacrum. Bats (for reasons that are unclear) fuse the skull but not other skeletal bones to the extent birds and pterosaurs do.

Avian notarium (fused back bones).
Which raises a question- what about the rest of the skeleton in marine animals? Do we get extra bones there, too?
It does seem that plesiosaurs and ichthyosaurs have a fairly high number of skeletal elements overall. The most striking example however is in the neck of elasmosaurs, which has a huge number of vertebrae. The back doesn’t really have that many by comparison. This suggests flexibility is another important determinant of where you get a lot of bones- it’s not just about weight reduction, but about what these bones do, and in some places it’s just useful to have more joints.
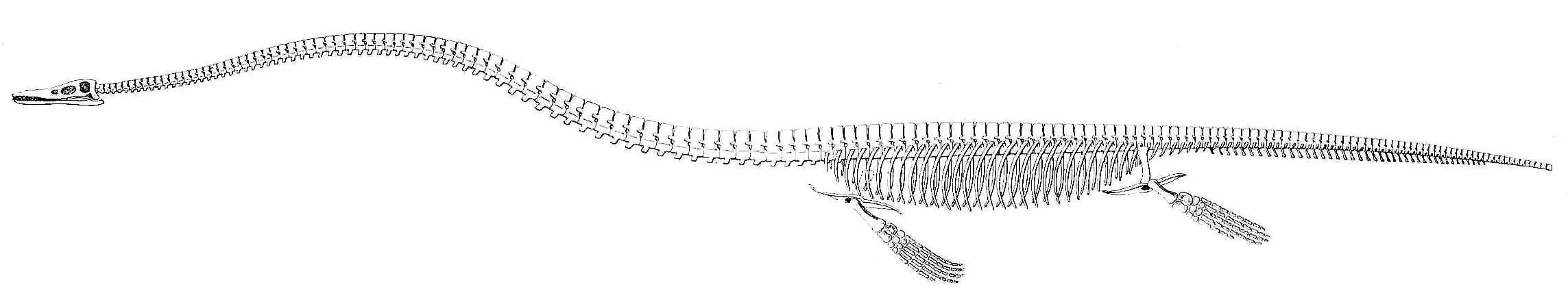
Elasmosaurus, showing the incredibly high number of neck vertebrae
There are also terrestrial animals with huge numbers of vertebrae- snakes - which suggests mobility is an important consideration, it’s not just about weight reduction. And then swans, which fly, can have 25 or more neck vertebrae.
A Unified Theory of Bone Number
The number of bones is heavily, maybe primarily driven by structural constraints. In retrospect this seems obvious. The function of the skeleton is structural, it acts to provide a combination of rigidity, and flexibility.
One of the biggest structural constraints is weight, or more specifically, strength to weight ratios. Moving from sea to land, the skeleton’s weight imposes a much higher cost, you have to carry every ounce of bone, so there’s a strong incentive to be more efficient. This means getting rid of bones, and fusing bones together. Flying animals have an even stronger pressure to reduce weight, and take this further.
Going the other direction releases this constraint: floating weightless or near-weightless in water there’s less pressure to get rid of extra elements and extra weight. Other design constraints become more important.
In certain contexts, having more bones and joints is advantageous. The high number of bones and knuckles in the flippers of marine tetrapods probably allows them to change shape subtly and precisely to maneuver and propel themselves through the water. Once plesiosaurs, mosasaurs and ichthyosaurs fully commit to life in water, and stop coming back to land to reproduce, this selective pressure takes over.
The final piece of the puzzle involves developmental constraints. Evolution works by tweaking the programs of our genetic code, mutation by mutation, and certain things are easier to change than others.
It seems to be easy to add bones to the fingers. Adding fingers is harder. And so is adding that little mystery bone in plesiosaurs, whatever it is: it only seems to evolve once, a unique one-off event in tetrapod evolution. The shared developmental patterns of humans, whales, plesiosaurs, ichthyosaurs, mosasaurs make it easier to evolve in certain directions, harder to evolve in another. There are rules. But those rules can be broken, sometimes. The one-two-many rule, for example, is very conservative, but not totally inviolable, as plesiosaurs show. Sometimes you think you know how evolution operates, and an animal comes along, and surprises you. Nature is funny that way.
These “rules” like the one-two-many rule exist because, due to shared ancestry, tetrapods have broadly similar developmental programs, genes that are turned on and off, upregulated and downregulated to make limbs, like a sort of computer program, except instead of rendering pixels and polygons on your screen, it draws in tissues like bone and cartilage.
All tetrapods inherited the same basic developmental program, which comes with limits. One is that it is easier to add minor tweaks at the very end of development than changes early on. Messing with the developmental program earlier is more likely to cause lethal mutations, since a mistake early on affects everything downstream- so maybe it’s easier to affect the number of finger bones (which form last) than the number of fingers (which is specified earlier), or the number of bones in the forelimb (which form even earlier). By analogy, making a mistake when you pour the foundation of your house is more likely to cause your building to have structural issues than making a mistake with the cornices, the attic, and the siding, since everything that comes later depends on the foundation being sound. A builder would tend to be very conservative in making changes to how he pours his foundations, but take more liberty with the later steps.
But the truth is, we have this very basic problem— aquatic animals tended to evolve lots of extra bones compared to their terrestrial relatives. And we can guess why this is, but we don’t really know for sure.
We don’t know why the plesiosaur Mystery Bone evolved, or how, or even what to call it.